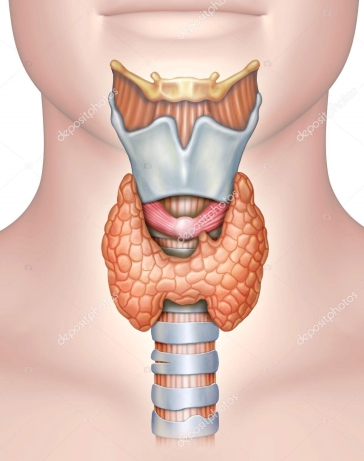
Como sabemos, la glándula tiroides es un órgano endocrino situado en el cuello. Su posición, cerca del cartílago tiroideo le proporciona el nombre a este órgano pues tiroides deriva de la palabra griega «thyros» que significa escudo, ya que originalmente se creía que protegía la laringe. Embriológicamente la tiroides se desarrolla en la base de la lengua de la fusión de tres estructuras y desciende desde donde su ubica durante la gestación a su posición final en la parte anterior del cuello.
ANATOMIA DE LA TIROIDES
Ubicación y aspectos generales
La glándula tiroides es una estructura impar, casi simétrica, situada adelante y a los lados de la tráquea y de la faringe. Está ubicada en la parte mediana del tercio inferior del cuello. Por su concavidad posterior, rodea el eje visceral aerodigestivo.

La tiroides del adulto es de color marrón y consistencia firme; se localiza detrás de los músculos esternotiroideo y esternohioideo. La glándula tiroides normal pesa alrededor de 20 g, pero su peso varia según sean el peso corporal y el consumo de yodo. Los lóbulos tiroideos están adyacentes a los cartílagos tiroides y unidos en la línea media por un istmo que habitualmente se encuentra justo debajo del cartílago cricoides.
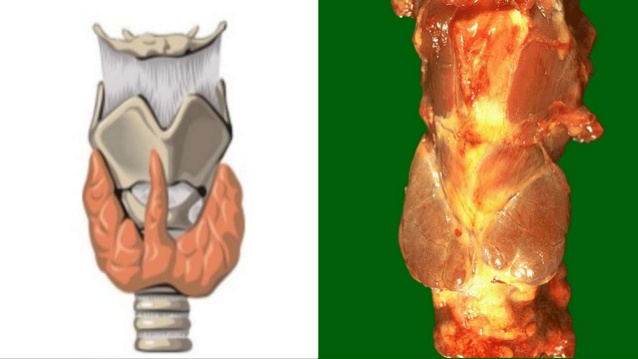
Configuración externa
La glándula tiroides está constituida por dos lóbulos reunidos por un istmo transversal. El conjunto adopta la forma de una H.
Lóbulos tiroideos
Se distinguen dos lóbulos tiroideos, derecho e izquierdo. Su parte inferior es más gruesa que la superior, que se adelgaza hacia arriba hasta terminar en un vértice. Se distinguen:
– Tres caras: anterolateral, convexa, superficial y cubierta por una capa muscular; posteromedial, cóncava, aplicada contra la cara anterolateral de la laringe y contra la tráquea, y en un plano profundo e inferior, en relación con la faringe y el esófago; posterior, la menos extensa, está orientada hacia atrás y algo lateralmente.
– Dos polos: inferior o base, convexo, orientado abajo y medialmente, que desciende hasta el 5° o 6° anillo de la tráquea; superior o vértice, más afinado, dirigido arriba y atrás, en contacto con el tercio inferior y lateral de la lámina del cartílago tiroides.
– Tres bordes: anteromedial, dirigido hacia abajo, adelante y medialmente; posteriores medial y lateral, redondeados, que reúnen insensiblemente las caras adyacentes.
En casi 50% de las personas existe un lóbulo piramidal. Los lóbulos tiroideos se extienden hacia el cartílago tiroides medial, por arriba, y yacen junto a las vainas carotideas y a los lados los músculos esternocleidomastoideos. Los músculos infrahioideos (esternohioideo, esternotiroideo y el vientre superior del omohioideo) se localizan por delante y reciben inervación del asa cervical (asa del hipogloso). La glándula tiroides está envuelta por una fascia laxa que se forma a partir de la división de la fascia cervical en sus partes anterior y posterior. La capsula verdadera de la tiroides es una capa fibrosa delgada muy adherente de la que se proyectan tabiques que penetran la glándula y forman seudolóbulos. La capsula tiroidea se condensa en el ligamento suspensorio posterior o de Berry cerca del cartílago cricoides y los anillos traqueales superiores.
ISTMO

Reúne a los dos lóbulos, con los cuales se continúa sin limite preciso. La cara anterior es convexa. La cara posterior, cóncava, abraza a los dos primeros cartílagos traqueales. Su borde inferior, corto y cóncavo hacia abajo, está a veces desarrollado en un lóbulo tiroideo medio. Su borde superior, cóncavo hacia arriba, llega cerca del cartílago cricoides. Desde la parte media (o ligeramente a la izquierda) del borde superior, asciende el lóbulo piramidal [pirámide de Lalouette], que es una prolongación aplanada, alargada, cuyo vértice afinado alcanza el borde superior del cartílago tiroides y puede llegar al hueso hioides, desde donde está prolongado por un «ligamento suspensor», vestigio del conducto tirogloso.
RELACIONES
La glándula tiroides está íntimamente rodeada por una cápsula fibrosa. Ésta se adhiere y envuelve al órgano, hacia cuyo interior emite prolongaciones. Por su intermedio contrae relaciones: adelante, con los planos musculares y fascias; atrás, con los elementos viscerales y vasculonerviosos.
Relaciones anterolaterales o superficiales
Corresponden a la cara anterior del istmo y anterolateral de los lóbulos, que por fuera de la cápsula fibrosa y de la vaina visceral están cubiertos sucesivamente por:
– La piel, el tejido subcutáneo y el músculo platisma.
– La hoja superficial de la fascia cervical, tendida entre ambos músculos esternocleidomastoideos que cubren la parte lateral de los lóbulos.
– Luego los músculos infrahioideos, reunidos por la hoja pretraqueal de la fascia cervical.
El músculo esternocleidomastoideo, superficial, transcurre oblicuo, hacia arriba y en sentido lateral. Más profundamente y en el plano de la hoja pretraqueal de la fascia
cervical, se encuentran el músculo omohioideo y el músculo esternohioideo. Más profundo, el músculo esternotiroideo, algo oblicuo hacia arriba y lateralmente. Ambos planos musculofasciales, separados y distintos, se adhieren en la línea media constituyendo la línea mediana infrahioidea.
Relaciones posteriores o profundas
También por fuera de la vaina visceral, se relacionan con el istmo y con las caras posterior y posteromedial de los lóbulos.
– En la línea mediana, el istmo corresponde por su concavidad a los cartílagos traqueales 2° y 3° (si se encuentra muy desarrollado, puede llegar hasta el cricoides). En su cara posterior se observa un plexo venoso profundo, origen de las venas tiroideas inferiores, que emergen por el borde inferior. Por este último penetra a la glándula la arteria tiroidea ima, cuando existe. El borde superior se relaciona con el espacio prelaríngeo que aloja ganglios prelaríngeos. De este borde superior se desprende el lóbulo piramidal.
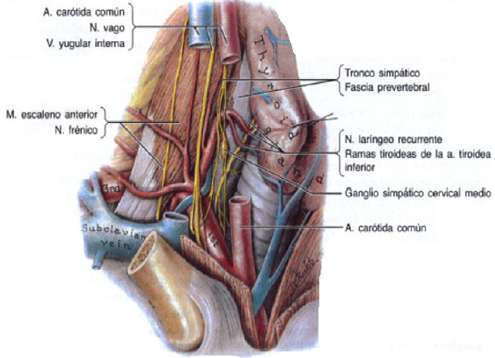
 – Lateralmente, la cara posterior de los lóbulos está delante del eje vasculonervioso yugulocarotídeo, con la arteria carótida común, medialmente, y la vena yugular interna, lateralmente. Entre ambas y atrás, se encuentra el nervio vago. El asa cervical se desliza con los nervios cardíacos cervicales superiores por delante de ella. Los ganglios linfáticos laterales del grupo cervical profundo son laterales a la vena yugular interna y el tronco simpático cervical es prevertebral.
– Lateralmente, la cara posterior de los lóbulos está delante del eje vasculonervioso yugulocarotídeo, con la arteria carótida común, medialmente, y la vena yugular interna, lateralmente. Entre ambas y atrás, se encuentra el nervio vago. El asa cervical se desliza con los nervios cardíacos cervicales superiores por delante de ella. Los ganglios linfáticos laterales del grupo cervical profundo son laterales a la vena yugular interna y el tronco simpático cervical es prevertebral.
La cara posteromedial de los lóbulos se amolda al conducto aerodigestivo; está aplicada a la laringofaringe arriba y a la tráquea y al esófago más abajo. El borde izquierdo de este último excede hacia lateral la cara posterior de la tráquea, con lo cual se encuentra en contado con el lóbulo izquierdo. Esta cara se relaciona con:
– Las glándulas paratiroides.
– La arteria tiroidea inferior.
IRRIGACIÓN
La irrigación de la glándula tiroides está dada por:
 Arterias tiroideas superiores
Arterias tiroideas superiores
• Nacen de las arterias carótidas externas ipsolaterales y se dividen en ramas anterior y posterior en el vértice de los lóbulos tiroideos.
Arterias tiroideas inferiores
• Provienen del tronco tirocervical, muy poco después de su origen en las arterias subclavias. Las arterias tiroideas inferiores se dirigen hacia arriba en el cuello, posteriores a la vaina carotidea, para entrar a los lóbulos tiroideos en su punto intermedio.
Arteria tiroidea media
• Nace de la aorta o el tronco braquiocefálico en 1 a 4% de las personas para entrar al istmo o sustituir a una arteria tiroidea inferior faltante. La arteria tiroidea inferior se cruza con el nervio laríngeo recurrente (RLN, recurrent laryngeal nerve), lo que obliga a identificar dicho nervio antes de ligar las ramas arteriales.


DRENAJE VENOSO
Se produce por múltiples venas superficiales pequeñas que confluyen para formar tres conjuntos de venas: las venas tiroideas superiores, medias e inferiores.
• Las primeras discurren con las arterias tiroideas en ambos lados.
• La vena o venas tiroideas medias son las menos constantes. Las venas superiores y medias drenan de manera directa en las venas yugulares internas; las venas tiroideas inferiores a menudo forman un plexo que drena en las venas braquiocefálicas.
INERVACIÓN
A lo largo de su trayecto en el cuello es posible que los RLN emitan ramas y pasen por delante o detrás o se intercalen con las ramas de la arteria tiroidea inferior.
El nervio laríngeo recurrente izquierdo (RLNI):
• procede del nervio vago en el sitio en que este cruza el cayado aórtico,
• forma un asa alrededor del ligamento arterioso y asciende por la parte medial en el cuello dentro de la hendidura traqueo esofágico.
El nervio laríngeo recurrente derecho (RLND):
• nace del N. vago en su cruce con la arteria subclavia derecha.
• pasa posterior a la arteria antes de ascender por el cuello.
• su trayecto es mas oblicuo que el del RLN izquierdo, no es recurrente en 0.5 a 1% de las personas y con frecuencia se relaciona con una anomalía vascular en esta situación.
Los RLN izquierdos solo se presentan en pacientes con situs inversus, y la paratiroides superior casi siempre está en un punto dorsal respecto del plano del nervio, mientras que la inferior es ventral en relación con el nervio, cayado aórtico derecho. Para identificar los nervios o sus ramas muchas veces es necesaria la movilización de la parte más lateral y posterior de la glándula tiroides, el tubérculo de Zuckerkandl, a nivel del cartílago cricoides. No es raro que los últimos segmentos de los nervios se proyecten hacia abajo del tubérculo muy cerca del ligamento de Berry. Algunos de los nervios laríngeos recurrentes terminan cuando entran a la laringe por detrás del musculo cricotiroideo.
Los RLN inervan todos los músculos intrínsecos de la laringe, excepto los músculos cricotiroideos que están inervados por los nervios laríngeos externos.
Nervios laríngeos superiores
• también se originan en los nervios vagos.
• origen en la base del cráneo,
• se proyectan a lo largo de la arteria carótida interna
• se dividen en dos ramas al nivel del hueso hioides.
• La rama interna del nervio laríngeo superior tiene función sensitiva para la laringe supraglótica. La lesión de este nervio es poco común en los procedimientos tiroideos, pero su ocurrencia puede ocasionar aspiración.
• La rama externa del nervio laríngeo superior se encuentra sobre el musculo constrictor laríngeo inferior y desciende junto al vaso tiroideo superior antes de inervar al musculo cricotiroideo.
HISTOLOGÍA DE LA TIROIDES
Por su parte externa, la glándula tiroides está recubierta por una delgada capa de tejido conectivo (una parte de la fascia visceral del cuello), separada por tejido conectivo laxo de una capa de tejido conectivo denso unida al tejido glandular, que forma una cápsula fibrosa verdadera a su alrededor. La existencia de una cápsula de dos capas tiene importancia quirúrgica práctica, dado que, en consecuencia, es más fácil aislar la glándula durante la cirugía.
Características histológicas de la glándula tiroides
La glándula tiroides se compone, sobre todo, de quistes (gr.kystis, vejiga) o folículos separados por escaso tejido conectivo interfolicular. Los folículos son la unidad estructural y funcional, y están compuestos por epitelio cúbico simple que rodea un espacio lleno de una sustancia viscosa, el coloide.
El tamaño de los folículos es variable, entre unos 50 pm y hasta1 mm de diámetro, por lo que los más grandes se ven a simple vista. Están rodeados por una delgada membrana basal y el tejido conectivo intersticial forma un reticulado a su alrededor.
El coloide es el producto de secreción de las células foliculares, que se almacena fuera de la célula. En los cortes histológicos comunes teñidos con HE, el coloide es eosinófilo franco. Se tiñe con intensidad con la reacción de PAS, dado que la tiroglobulina, el componente predominante del coloide, es una glucoproteína. La tiroglobulina contiene precursores de tiroxina y triyodotironina, formados por yodización y acoplamiento de a pares de restos de tirosina en la cadena peptídica de la tiroglobulina. Los folículos tiroideos están compuestos por:
Células foliculares
Por lo general son cúbicas, pero hay notable variación de altura, de acuerdo con el estado funcional de la glándula. Cuando la tiroides presenta una relativa inactividad, las células son casi planas, mientras que son cilíndricas altas en la glándula con gran actividad. Sin embargo, no se puede evaluar el estado funcional glandular sólo sobre la base de la altura del epitelio.
Todas las células foliculares limitan con la luz y poseen un núcleo redondo bastante claro (en las células foliculares aplanadas, el núcleo también es plano). El citoplasma presenta basofilia moderada y contiene vesículas apicales, que se tiñen de modo similar al coloide de la luz folicular.
Con microscopia electrónica se observa un retículo endoplasmático rugoso bien desarrollado, sobre todo en dirección basal y lateral al núcleo. El aparato de Golgi se localiza en posición supranuclear, mientras que las mitocondrias están dispersas por todo el citoplasma. En el citoplasma apical, se distinguen tres tipos de gránulos o vesículas, todas limitadas por membrana:
1) vesículas pequeñas, no muy electrodensas, con un diámetro de unos 200 nm, denominadas gránulos subapicales, y que contienen tiroglobulina para liberar en la luz folicular por exocitosis.
2) Vesículas más grandes, con un diámetro de hasta 4 flm, llamadas gotas de coloide, idénticas a las vesículas apicales visibles con microscopio óptico. Las gotas de coloide contienen coloide captado desde la luz por endocitosis.
3) Por último, se distinguen vesículas muy densas que son Iisosomas primarios; a menudo se observan en proceso de fusión con las gotas de coloide.
La superficie luminal posee microvellosidades cortas, y a menudo se observan seudópodos que rodean las gotas de coloide durante la endocitosis, sobre todo ante una fuerte estimulación de la glándula por TSH. En la superficie celular lateral es característico encontrar complejos de unión yuxtaluminales. Las células foliculares sintetizan, almacenan y secretan las hormonas triyodotironina (T3) y tiroxina (tetrayodotironina, T4) ambas formadas por dos restos yodados del aminoácido tirosina. El ciclo secretor de estas hormonas se caracteriza porque la síntesis es parcialmente extracelular y el almacenamiento lo es en su totalidad, y tiene lugar en la luz folicular.
La síntesis de tiroglobulina ocurre en el retículo endoplasmático rugoso y en el aparato de Golgi. La tiroglobulina es una glucoproteína en la que alrededor del 3% de la proteína se compone de restos de tirosina, unos 100 por molécula. La parte proteica es sintetizada por el retículo endoplasmático rugoso, donde también se le adiciona parte de los hidratos de carbono.
En el aparato de Golgi se agrega el resto de los hidratos de carbono y el producto de secreción es liberado en los gránulos subapicales. La enzima peroxidasa tiroidea también es sintetizada en el retículo endoplasmático rugoso e incorporada a la membrana de las vesículas subapicales que, en consecuencia, transportan tiroglobulina y peroxidasa tiroidea. Los gránulos subapicales migran a la superficie celular apical y liberan la tiroglobulina a la luz folicular por exocitosis. La fusión de la membrana de los gránulos apicales con la membrana celular incorpora la peroxidasa tiroidea en la membrana celular apical.
La yodación de la tiroglobulina tiene lugar en la parte periférica de la luz folicular. La glándula tiroides es capaz de concentrar iones de yodo inorgánico (yoduro) del plasma por un mecanismo de transporte activo a través de la membrana celular basal. La captación ocurre por simporte con iones sodio y es mediada indirectamente por la ATPasa de Na+-K+, también localizada en el plasmalema basal. Los iones yodo migran a través de la célula y son liberados en la luz folicular desde la región celular apical, posiblemente a través de un canal de yoduro transmembrana. En la luz folicular cerca de la superficie celular apical, los iones yodo captados sufren primero una oxidación catalizada por la enzima peroxidasa tiroidea antes de ser incorporados en la tiroglobulina.
Con esta incorporación, se yodan menos de 20 restos de tirosina en cada molécula de tiroglobulina por un proceso que también es catalizado por la peroxidasa tiroidea y tiene lugar en la luz folicular en estrecha relación con la superficie celular apical. Uno o dos átomos de yodo se fijan a un resto de tirosina, por lo que se forma MIT (ing. monoiodotyrosine) y DIT (ing. diiodotyrosine). Las hormonas tiroideas se forman por acoplamiento de una MIT a una DIT (T3) o de dos DIT entre sí (T4). Este acoplamiento puede ocurrir cuando MIT y DIT están muy cercanas en la molécula de tiroglobulina. Despué s del acoplamiento, T, y T4 permanecen en la molécula de tiroglobulina y se ubican en la luz del folículo.
s del acoplamiento, T, y T4 permanecen en la molécula de tiroglobulina y se ubican en la luz del folículo.
Como se mencionó, la captación de coloide tiene lugar por endocitosis, con formación de las gotas de coloide apicales que se fusionan con lisosomas primarios, cuyas enzimas después escinden T3 y T4. Las hormonas abandonan entonces la célula a través de la membrana celular basal y llegan al torrente sanguíneo. Se libera aproximadamente 20 veces más T4 que T3.
Células C
Estas células se encuentran en número mucho menor que las foliculares. Es característica su localización basal al epitelio folicular y nunca están en contacto con la luz. Se encuentran aisladas o en grupos pequeños de 3-4 células, siempre ubicadas dentro de la membrana basal, es decir, entre ésta y las células foliculares. Las células C son ovaladas y más grandes y claras que las células foliculares; además, poseen un núcleo más grande y a menudo más claro. Son difíciles de distinguir en los preparados comunes teñidos con HE, pero pueden demostrarse mediante técnicas de impregnación argéntica o inmunohistoquímicas con anticuerpos contra el producto de secreción calcitonina.
Con microscopia electrónica, se observa un retículo endoplasmático rugoso y un complejo de Golgi bien desarrollados. El citoplasma también contiene numerosas vesículas electrodensas limitadas por membrana y que contienen calcitonina. Al parecer, las células C no forman complejos de unión extensos con las células foliculares o entre sí, aunque se demostró la presencia de desmosomas entre las células C. La ausencia de unión con las células foliculares debe analizarse teniendo en cuenta que las células C se desarrollaron a partir de la cresta neural y se consideran parte del sistema neuroendocrino difuso.
Como se mencionó antes, las células C producen la hormona calcitonina, un polipéptido contenido en vesículas de secreción que se liberan por exocitosis. La calcitonina disminuye la concentración sanguínea de calcio al inhibir la resorción de tejido óseo. Esto ocurre por inhibición de la actividad de los osteoclastos (que poseen receptores para calcitonina), por lo que disminuye la velocidad de remodelación del tejido óseo.
La secreción de calcitonina es regulada por la concentración sanguínea de calcio por retrocontrol negativo. Así, un aumento de la concentración de calcio en sangre produce mayor liberación de calcitonina por las células C; esto induce menor resorción ósea, lo cual causa descenso de la concentración de calcio en la sangre. Al parecer, la hormona paratiroidea tiene mucha mayor importancia para la regulación normal de la homeostasis del calcio.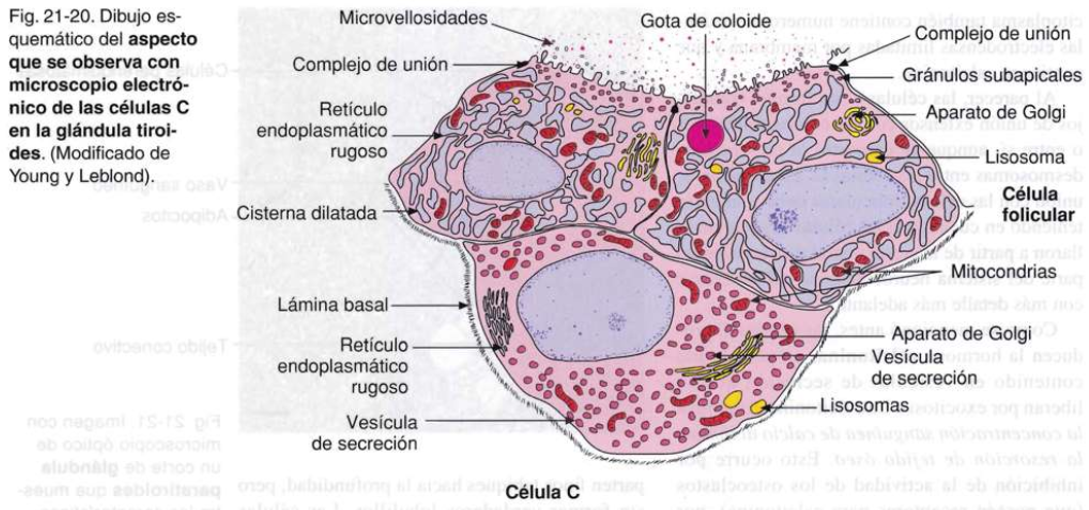
FISIOLOGÍA DE LA GLÁNDULA TIROIDES
Metabolismo del yodo.
Las necesidades diarias promedio de yodo son de 0.1 mg, que puede obtenerse de alimentos como pescado, leche y huevos, aditivos del pan o la sal. En el estómago y el yeyuno, el yodo se convierte pronto en yoduro y se absorbe a la corriente sanguínea, desde donde se distribuye de manera uniforme por todo el espacio extracelular.
Síntesis de hormonas
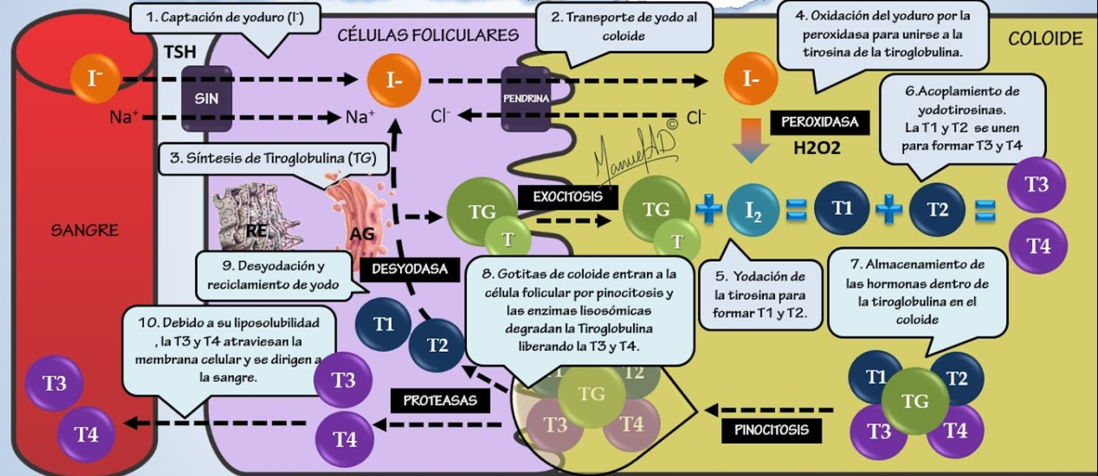
El yoduro se transporta por mecanismos activos hacia el interior de las células foliculares mediante un proceso dependiente de trifosfato de adenosina (ATP). La tiroides es el sitio de almacenamiento de más del 90% del yodo del cuerpo y representa un tercio de la pérdida plasmática del mismo. El yodo plasmático restante se excreta por vía renal. Dichos mecanismos corresponden: Síntesis, secreción y transporte de hormona tiroidea. La síntesis de hormona tiroidea incluye varios pasos:
- Primero, atrapamiento del yodo, implica el transporte activo (dependiente de ATP) del yodo a través de la membrana basal del tirocito mediante una proteína intrínseca de la membrana, el cotransportador unidireccional Na+/I−. La tiroglobulina (Tg) es una glucoproteína grande (660 kDa) que se encuentra en los folículos tiroideos y tiene cuatro residuos de tirosilo.
- El segundo paso en la síntesis hormonal supone la oxidación de yodo a yoduro y la yodación de los residuos de tirosina de la tiroglobulina para formar monoyodotirosina (MIT) y diyodotirosina (DIT). La peroxidasa tiroidea cataliza ambos procesos. Se cree que la proteína pendrina, recién identificada, media la salida de yodo en la membrana apical.
- El tercer paso conduce al acoplamiento de dos moléculas de DIT para formar la tetrayodotironina o tiroxina (T4) y una molécula de DIT con una de MIT para formar la 3,5,3′-triyodotironina (T3) o la 3,3′,5′-triyodotironina inversa (rT3). Cuando reciben el estimulo de la TSH, los tirocitos forman seudópodos que rodean porciones de la membrana celular que contienen tiroglobulina, la que a su vez se fusiona con lisosomas que poseen enzimas.
- El cuarto paso, la tiroglobulina se hidroliza para liberar yodotironinas libres (T3 y T4), además de monotirosinas y diyodotirosinas.
Estas últimas se separan del yodo en el quinto paso para producir yodo libre, que el tirocito utiliza de nueva cuenta. En el estado eutiroideo, T4 se produce y libera de la glándula tiroides, pero solo 20% del total de T3 se elabora en la tiroides. La mayor parte de la T3 se crea por desyodación periférica (remoción de 5′-yodo del anillo exterior) de T4 en el hígado, musculo, riñones e hipófisis anterior, reacción que cataliza la 5′-monodesyodinasa. Parte de la T4 se convierte en rT3, compuesto sin actividad metabólica, por desyodación del anillo interior de T4.
En trastornos como la enfermedad de Graves, el bocio tóxico multinodular o una glándula tiroides estimulada, la proporción de T3 liberada de la glándula puede elevarse en grado notorio.
Las hormonas tiroideas se transportan en el suero unidas con proteínas transportadoras como la globulina de unión con tiroxina (TBG), la prealbúmina de unión con tiroxina y la albumina. Solo una pequeña fracción (0.02%) de la hormona tiroidea (T3 y T4) está libre (no unida) y es el componente con actividad fisiológica. La T3 es la más potente de las dos hormonas tiroideas, aunque su concentración plasmática es mucho menor a la de T4. La T3 se une con menor fuerza a la proteína en el plasma que T4 y también entra a los tejidos con mayor facilidad. La T3 posee una actividad tres a cuatro veces mayor que T4 por unidad de peso, con una vida media aproximada de un día, en comparación con los casi siete días de la tiroxina.
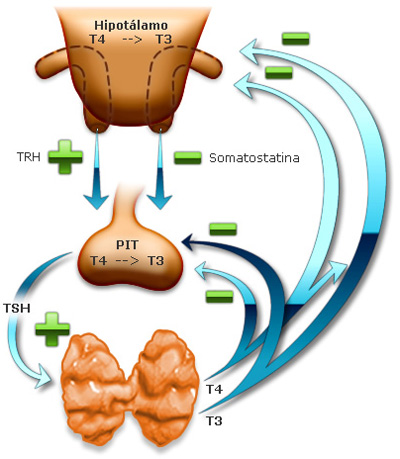 El eje hipotálamo-hipófisis-tiroides controla la secreción de hormona tiroidea. El hipotálamo produce un péptido, la hormona liberadora de tirotropína (TRH, thyrotropin-releasing hormone), que estimula a la hipófisis para que libere TSH o tiroestimulante. La TRH llega a la hipófisis en la circulación venosa portal. La TSH es un glucopeptido de 28 kDa que media el atrapamiento de yodo, la secreción y liberación de hormonas tiroideas, además de aumentar la celularidad y vascularidad de la glándula tiroides. El receptor para TSH pertenece a una familia de receptores unidos con proteína G, tiene siete dominios transmembrana y utiliza cAMP como vía para la transducción de señal.
El eje hipotálamo-hipófisis-tiroides controla la secreción de hormona tiroidea. El hipotálamo produce un péptido, la hormona liberadora de tirotropína (TRH, thyrotropin-releasing hormone), que estimula a la hipófisis para que libere TSH o tiroestimulante. La TRH llega a la hipófisis en la circulación venosa portal. La TSH es un glucopeptido de 28 kDa que media el atrapamiento de yodo, la secreción y liberación de hormonas tiroideas, además de aumentar la celularidad y vascularidad de la glándula tiroides. El receptor para TSH pertenece a una familia de receptores unidos con proteína G, tiene siete dominios transmembrana y utiliza cAMP como vía para la transducción de señal.
La secreción de TSH en la hipófisis anterior también la regula un ciclo de retroalimentación negativa por la T4 y la T3. Como la hipófisis tiene la capacidad de convertir T4 en T3, se cree que esta última es más importante para el control por retroalimentación. La T3 también inhibe la liberación de TRH. La glándula tiroides también posee la capacidad de autorregulación, lo cual le permite modificar su función de manera independiente de las concentraciones de TSH. Como adaptación al bajo consumo de yodo, la glándula sintetiza en forma preferente T3 en lugar de T4, lo que aumenta la eficiencia de la hormona secretada.
En el caso de exceso de yodo, se suprimen el transporte de éste, la generación de peróxido, la síntesis y la secreción de hormonas tiroideas. Las dosis excesivas de yodo pueden precipitar un incremento inicial en la urbanificación seguida de supresión, un fenómeno llamado efecto de Wolff-Chaikoff. La adrenalina y la gonadotropina coriónica humana (hCG) estimulan la producción de hormona tiroidea. Por lo tanto, durante el embarazo se registran concentraciones elevadas de hormona tiroidea, así como en todos los tumores malignos ginecológicos, como la mola hidatidiforme. En cambio, los glucocorticoides inhiben la producción de hormona tiroidea. En los pacientes graves puede haber reducción de las hormonas tiroideas periféricas, sin aumento compensatorio de las concentraciones de TSH, lo que causa síndrome de enfermedad eutiroidea.
Funciones de las hormonas tiroideas
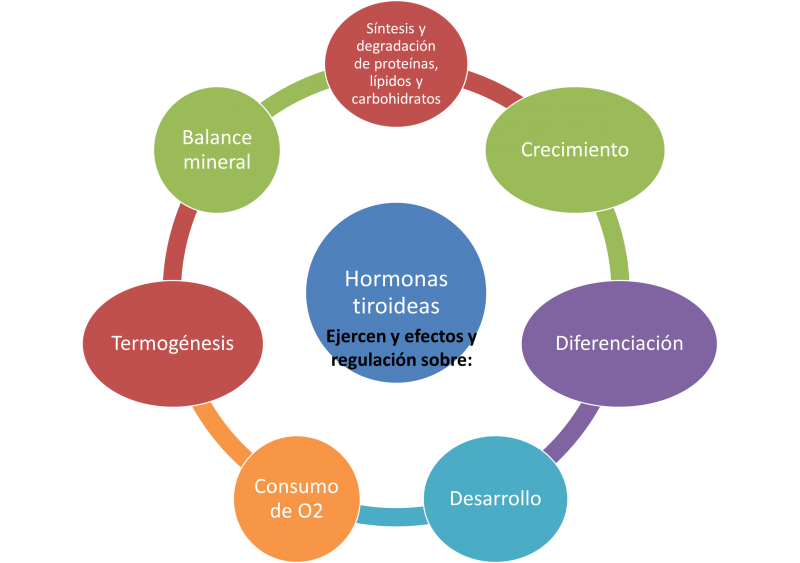
La hormona tiroidea libre penetra la membrana celular por difusión o mediante transportadores específicos y llega hasta la membrana nuclear mediante la unión con proteínas específicas. La T4 pierde yodo para formar T3 y entra mediante transporte activo al núcleo, donde se une con el receptor para la hormona tiroidea. El receptor para T3 es similar a los receptores nucleares para glucocorticoides, mineralocorticoides, estrógenos, vitamina D y acido retinoico. En los seres humanos, dos tipos de genes para receptor de T3 (α y β) se localizan en los cromosomas 3 y 17. La expresión del receptor tiroideo depende de las concentraciones periféricas de hormonas tiroideas y es específica para cada tejido: la forma α abunda en el sistema nervioso central, en tanto que la forma β predomina en el hígado.
Cada producto genético tiene un dominio amino terminal independiente del ligando; un dominio carboxilo terminal para unión con ligando y regiones centrales para unión con DNA. La unión de la hormona tiroidea induce la transcripción y traducción de genes con respuesta especifica a la hormona. Las hormonas tiroideas afectan a casi todos los sistemas del cuerpo. Son importantes para el desarrollo cerebral y la maduración esquelética del feto. La T3 eleva el consumo de oxigeno, la tasa metabólica basal y la producción de calor mediante la estimulación de la ATP-asa de Na+/K+ en varios tejidos. También tiene un efecto inotrópico y cronotrópico positivo sobre el corazón, porque aumenta la transcripción de la ATP-asa de calcio en el retículo sarcoplásmico e incrementa las concentraciones de receptores adrenérgicos β y las de proteínas G. Los receptores miocárdicos α disminuyen y las acciones de las catecolaminas se amplifican. Las hormonas tiroideas son las que mantienen el estimulo normal a la hipoxia e hipercapnia en el centro respiratorio del cerebro. También incrementan la motilidad gastrointestinal, lo que produce diarrea en el hipertiroidismo y estreñimiento en el hipotiroidismo; asimismo, aumentan el recambio óseo y proteínico, así como la velocidad de la contracción y relajación muscular. De igual modo, intensifican la glucogenólisis, la gluconeogénesis hepática, la absorción intestinal de glucosa, y la síntesis y degradación del colesterol.
BIBLIOGRAFÍA
*Latarjet-Ruiz Liard. Anatomía Humana. Editorial Panamericana. IV Edición
*Finn Geneser. Histología. Editorial Panamericana. III Edición
*Guyton-Hall. Tratado de Fisiología Médica. ELSERVIER SAUNDERS. XII Edición
